
We use cookies to ensure our website works properly and to personalise your experience. Cookies policy
Women's College of Pharmacy Peth Vadgaon
The zoonotic pathogen Nipah virus (NiV) has emerged as a significant global health threat, provoking outbreaks characterised by high mortality and substantial transmission potential. First identified in 1998?1999 in Malaysia and Singapore in association with pig farms and fruit bat reservoirs, NiV is now recognised as a member of the genus Henipavirus within the family Paramyxoviridae. This review article provides a comprehensive synthesis of current knowledge on NiV, including its epidemiology, molecular virology, clinical features, diagnostic approaches, therapeutic strategies, and public health management. We examine outbreak patterns in Southeast Asia, assessing reservoir hosts (predominantly Pteropus spp. fruit bats), intermediate animal hosts (notably pigs), and documented human-to-human transmission events. In terms of virology, NiV possesses a negative-sense single-stranded RNA genome (~18 kb) encoding six major structural proteins (N, P, M, F, G, and L). The viral glycoprotein fusion complex interacts with Ephrin-B2/B3 cellular receptors to mediate entry, enabling broad tissue tropism including endothelial and neuronal cells. The pathophysiological cascade involves endothelial damage, vasculitis, micro-infarction, blood?brain barrier penetration, and fulminant encephalitis or respiratory failure. Clinically, incubation periods of 5?14 days (occasionally longer) precede prodromal flu-like symptoms, followed by neurological decline, seizures, and acute respiratory distress in severe cases; relapsing or delayed-onset encephalitis has also been described. Diagnostic modalities include real-time RT-PCR, serology (IgM/IgG ELISA), virus isolation (BSL-4), and newer point-of-care assays under development. Management remains largely supportive, as there is currently no licensed antiviral or vaccine for NiV. Experimental therapeutics such as ribavirin, favipiravir, monoclonal antibodies (e.g., m102.4), and viral-vectored vaccine candidates are in varying phases of research. From a public health perspective, the adoption of One Health approaches, rigorous surveillance, bat and livestock monitoring, infection control in healthcare settings, and public education are critical to prevention. Recent outbreaks in India (particularly the state of Kerala in 2018 and 2023) and Bangladesh demonstrate both the ongoing threat and the effectiveness of rapid containment when robust systems are in place. Future challenges include elucidating spill-over mechanisms, refining diagnostics and therapeutics, and developing broadly protective vaccines. In conclusion, given the high case-fatality rate, propensity for human transmission, and absence of licenced countermeasures, NiV demands sustained global vigilance, research investment, and integrated human-animal-environmental health strategies.
The Nipah virus (NiV) is an enveloped, negative-sense RNA virus of the genus Henipavirus and family Paramyxoviridae. It emerged as a zoonotic pathogen of grave concern when it was first identified in the 1998-1999 outbreak in Malaysia and Singapore, in which pig farms and the fruit-bat reservoir were implicated (Chua et al., 2000; Luby & Gurley, 2012). The public health significance of NiV arises from its high case-fatality rate ranging from approximately 40 % to as much as 75 % or more in certain outbreaks (Luby et al., 2009; Hsu et al., 2004) its documented capacity for human-to-human transmission, and its classification by the World Health Organization (WHO) as a priority disease under the R&D Blueprint for emerging pathogens (WHO, 2023). Outbreaks occurring nearly annually in South and Southeast Asia (notably Bangladesh, India, Malaysia, and the Philippines) highlight the potential for wider spread, especially given bat reservoirs and increasing ecological interface between wildlife, livestock, and humans (Rahman et al., 2012; Lo & Rota, 2018).
This review article aims to provide a scholarly, evidence-based synthesis of current knowledge on NiV. Specifically, we examine:
(1) The historical discovery and major outbreaks of niv
(2) The virology and molecular structure of the virus
(3) The epidemiology of niv including global distribution, outbreak drivers, and transmission dynamics
(4) The pathophysiology of infection and organ involvement
(5) Clinical manifestations and diagnostic modalities
(6) Therapeutic strategies and vaccine development
(7) Public health measures and preventive efforts
(8) Recent outbreak experiences and lessons learned
(9) Future directions and remaining challenges; and
(10) In conclusion, the imperative for integrated control and research.
By synthesising findings from peer-reviewed literature (2010–2025) and major surveillance reports, this review seeks to inform clinicians, public health professionals, virologists, and policy-makers about the critical aspects of NiV and the necessary preparations for future emergence.
2. History and Discovery
The first recognized outbreak of Nipah virus occurred between September 1998 and May 1999 in Malaysia and Singapore, centred on pig-farm communities. The index cluster in Malaysia involved 265 human cases and 108 deaths, with a case-fatality rate (CFR) of approximately 40.8 % (Chua et al., 2000; WHO, 2001). The investigation revealed that domestic pigs acted as an amplifying intermediate host, with the reservoir implicated as fruit bats (genus Pteropus) that contaminated pig feed or housing with excreta (Chua et al., 2002). The outbreak spread to Singapore via pig imports, prompting > 1 million pigs to be culled. Following this initial outbreak, subsequent NiV outbreaks have occurred in Bangladesh since 2001, with nearly annual seasonal events (Luby et al., 2009; Hossain et al., 2008). The first major Indian outbreak occurred in Siliguri, West Bengal, in 2001 (Masrour et al., 2001), followed by another in Nadia district in 2007 (Gurley et al., 2007). In 2018, the state of Kerala, India, reported 23 confirmed and probable cases (17 deaths, CFR ~74 %) in May–June 2018, marking the first southern India outbreak (Joseph et al., 2019; Recurrent … Kerala, 2023). Additional outbreaks have occurred in 2021, 2023, 2024 and 2025 in Kerala, reaffirming the region as a hotspot. Key milestones in NiV research include: the identification of the virus in pigs and bats in Malaysia (Chua et al., 2002); recognition of human-to-human transmission in Bangladesh (Gurley et al., 2007; ICDDR, B, 2008); detailed elucidation of viral receptor interactions (Bonaparte et al., 2005); development of monoclonal antibodies and animal models (Bossart et al., 2008); and recommendation of NiV as a priority pathogen by WHO (WHO, 2015). Over the past two decades, NiV has transitioned from a primarily zoonotic pig-borne event to a pathogen with documented human-to-human and nosocomial transmission, thereby elevating its pandemic potential (Ang et al., 2018). In summary, the discovery and historical outbreak chronology of NiV depict an evolving zoonosis with significant human health implications, necessitating continuous vigilance and research.
3. Virology and Molecular Structure
The Nipah virus is classified as genus Henipavirus, family Paramyxoviridae. It is closely related to the Hendra virus (HeV) and shares features with paramyxoviruses such as measles and mumps viruses, though with notable differences in genome length and host range (Wang et al., 2003). NiV is an enveloped virus possessing a non-segmented, negative-sense single-stranded RNA genome of approximately 18 kb in length, which encodes six major structural proteins: nucleoprotein (N), phosphoprotein (P), matrix protein (M), fusion protein (F), attachment glycoprotein (G), and the large polymerase (L) (Diederich et al., 2016).
3.1. Genome organisation and proteins
3.2. Mechanism of cell entry and replication
NiV utilises the cellular receptors Ephrin-B2 and Ephrin-B3 on host cells (Bonaparte et al., 2005; Negrete et al., 2006). These receptors are highly conserved across mammalian species and are abundantly expressed in endothelial and neuronal tissues, explaining the virus’s broad tropism and severe neuropathology. The G protein binds Ephrin-B2/B3, triggering conformational changes that activate F protein, resulting in fusion of the viral and host membranes and release of the ribonucleoprotein complex into the cytoplasm. Viral replication proceeds via the polymerase L complex and produces mRNAs, antigenome, and progeny genomes; subsequent assembly at the plasma membrane involves M protein and budding of new virions.
3.3. Comparison with related viruses
Although measles virus and mumps virus (Paramyxoviridae) also use F and H/HN glycoproteins for entry, NiV has a broader host range and higher pathogenicity. In contrast to HeV, which primarily infects equines and humans (Australia), NiV has a broader zoonotic spectrum and more frequent human-to-human transmission. Genome variation studies indicate two major lineages of NiV: the Malaysia strain and the Bangladesh strain, with the latter associated with higher mortality and more frequent human-to-human spread (Luby et al., 2009).
3.4. Mutational diversity and implications for virulence
Although henipaviruses have relatively stable genomes compared to influenza or HIV, some sequence variation has been identified between outbreak strains. The differences in non-structural proteins (V, W) may influence interferon antagonism and thus virulence (Lo & Rota, 2018). Genomic surveillance (Twenty-five years review) has documented 754 human NiV cases and 435 deaths globally (CFR ~58 %) but also highlighted limited mutational drift to date (Twenty-five years … 2024). From a public-health perspective, the relative genome stability is reassuring for vaccine and therapeutic design, but the capacity for adaptation remains a concern. In sum, the molecular biology of NiV offers insight into its pathogenic potential (broad receptor usage, tissue tropism, immune evasion mechanisms), and underpins the rationale for antiviral and vaccine development.
4. Epidemiology
4.1 Global Distribution and Outbreak Patterns
The global distribution of NiV is predominantly within South and Southeast Asia, with spillover risk zones overlapping with the range of Pteropus spp. fruit bats. The first large outbreak occurred in Malaysia and Singapore in 1998–1999, with 265 human cases and 108 deaths. In Bangladesh, outbreaks have been recorded almost annually since 2001, often in the “date-palm sap” season (Luby et al., 2009; Hossain et al., 2008). In India, major outbreaks occurred in Siliguri (2001), Nadia (2007), and Kerala (2018, 2021, 2023–25) with variable case-fatality rates (Joseph et al., 2019; WHO 2025). Recently, the Philippines reported a NiV or NiV-like henipavirus outbreak (Philippines, 2014) (Gamage et al., 2017). According to a recent review, 754 human NiV cases with 435 deaths (CFR ≈ 58%) were reported globally over 25 years. Table 1 outlines a summary timeline of major outbreaks (country, year, CFR) for reference.
|
Country |
Year(s) |
Approximate Case-Fatality Rate |
|
Malaysia / Singapore |
1998–1999 |
~40 % (108/265) (Chua et al., 2000) |
|
Bangladesh |
2001–present |
~70–90 % in many outbreaks (Luby et al., 2009; Tackling NiV surveillance … 2021) |
|
India – Siliguri |
2001 |
~68 % (45 deaths) |
|
India – Kerala |
2018 |
~74 % (17/23) (Joseph et al., 2019) |
|
India – Kerala |
2023–25 |
Variable (e.g., 2023: 6 cases, 2 deaths, CFR ~33 %) |
Reservoir hosts are fruit bats of genus Pteropus (Pteropus vampyrus, P. giganteus, P. medius) which carry NiV asymptomatically and can shed virus via urine, saliva, or partially eaten fruit (Wacharapluesadee et al., 2010). Intermediate hosts have included pigs (Malaysia), and in some outbreaks direct bat-to-human or human-to-human transmission has occurred (Hossain et al., 2008; Gurley et al., 2007). Transmission modes include zoonotic (animal-to-human), human-to-human (direct contact, nosocomial), and foodborne (date-palm sap contaminated by bats) (Luby & Gurley, 2012). The geographical risk zones include Bangladesh, eastern India, southern India (Kerala), Malaysia, and potentially neighbouring countries where Pteropus bats are present (Gov.UK Nipah guidance 2024).
4.2 Factors Influencing Outbreaks
Environmental and ecological determinants play a major role in NiV spill-over. Deforestation, urban expansion, agricultural intensification, and wildlife habitat fragmentation increase human–bat–livestock contact (Field et al., 2015). Climate factors (particularly drinking of raw date-palm sap in winter months) facilitate bat access to sap containers, thereby increasing foodborne transmission (Rahman et al., 2012). Cultural practices such as consumption of raw date-palm sap in Bangladesh and India have been directly implicated (Luby et al., 2009). Livestock farming practices (pig density, proximity to bat roosts) are significant; the pig-to-human amplification seen in Malaysia underscores this point (Chua et al., 2002). Healthcare-associated transmission is further influenced by inadequate infection prevention and control (IPC) measures, overcrowded hospitals, delayed diagnosis, and exposure of healthcare workers observed in Bangladesh and Kerala outbreaks (Joseph et al., 2019).
4.3 Transmission Dynamics
The transmission dynamics of NiV illustrate its capacity for zoonotic spill-over and limited but significant human-to-human spread.
4.3.1. Reservoir-to-human transmission
Fruit bats (genus Pteropus) serve as the known natural reservoir of NiV. Humans may be infected through contact with bat urine, saliva, partially eaten fruit or contaminated date-palm sap. In Bangladesh, drinking raw date-palm sap contaminated by bat excreta is a well-documented mode of zoonotic transmission (Luby et al., 2009). In Malaysia, contamination of pig feed or stabling by bats was implicated (Chua et al., 2002). Transmission from bats to humans remains sporadic, but the risk persists in areas with bat–human interface.
4.3.2. Animal-to-human (intermediate host) transmission
Amplification of NiV in intermediate animal hosts increases human risk. In Malaysia, pigs acted as amplifying hosts: infected by bats, then transmitting NiV to farm workers with high virus loads and respiratory shedding (Chua et al., 2002). Similar events have not been widely documented elsewhere, but remain biologically plausible for other livestock species.
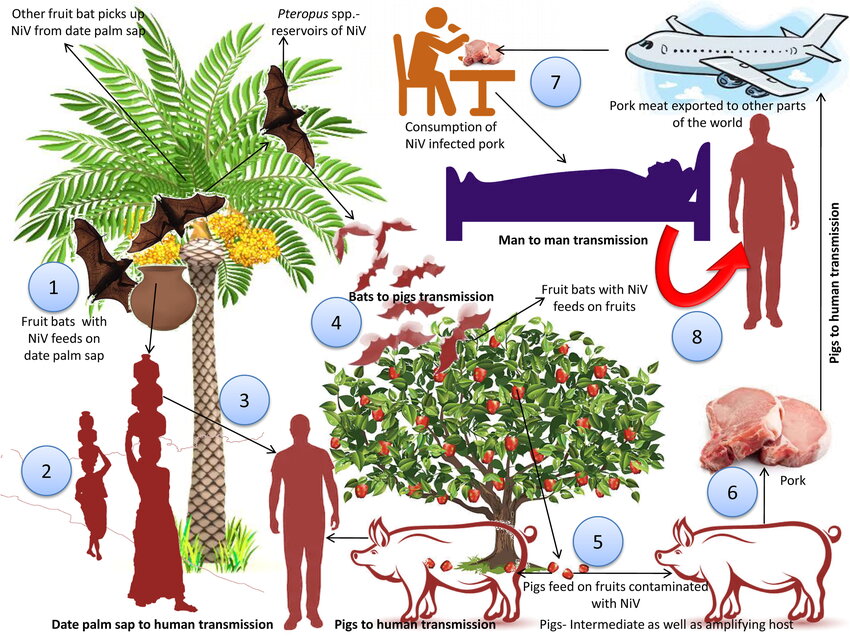
Fig no. 1: Schematic of NiV transmission cycle
4.3.3. Human-to-human and nosocomial transmission
Unlike many zoonoses, NiV has demonstrated human-to-human transmission, particularly in settings with close contact and respiratory involvement. In Bangladesh, studies of cases from 2001-2014 documented 14 years of investigations, revealing chains of transmission among caregivers and hospital personnel (N Engl J Med Transmission study). In Kerala (2018) the index case transmitted infection to a nurse and multiple other contacts in hospital setting (Joseph et al., 2019). Super-spreader events and nosocomial spread are therefore real concerns. However, the basic reproduction number (R?) for NiV remains below that of pathogens such as measles; modeling studies suggest limited sustained human-to-human transmission (Modelling NiV dynamics 2020).
4.3.4. Shedding, incubation and infectivity
Reported incubation periods of NiV range from 5–14 days, with some documented up to 45 days in rare cases (Wikipedia summary). Viral shedding in humans may occur in respiratory secretions, urine, and blood. The duration of shedding and infectivity are not fully characterised, but longer shedding in respiratory secretions may correlate with transmission risk (Hassan et al., 2020). Understanding of environmental survival (on fomites, in sap) remains incomplete. In summary, the transmission dynamics of NiV encompass zoonotic spill-over, animal-to-human amplification, and human-to-human transmission under facilitating conditions. While the R? remains modest, the high mortality and outbreak potential elevate NiV to a pathogen of concern.
5. Pathophysiology
The pathophysiology of NiV infection reflects its broad cellular tropism, potent endothelial and neuronal involvement, and capacity for multisystem disease.
5.1. Viral entry and tissue tropism
NiV binds to cellular receptors Ephrin-B2 and Ephrin-B3, which are expressed on endothelial cells, neurons, smooth muscle cells, and certain epithelial tissues (Bonaparte et al., 2005). This receptor distribution (especially in brainstem, lung, vasculature) explains the neurotropic and vasculitic manifestations of NiV. Following binding, membrane fusion mediated by the F glycoprotein facilitates viral entry. Once inside, NiV replicates in the cytoplasm, assembles new virions, and budding occurs at the plasma membrane. Endothelial infection leads to vascular injury, micro-infarcts, and haemorrhagic lesions.
5.2. Mechanism of disease progression
NiV primarily causes severe disease via a cascade of vascular and neurological injury:
5.3. Neuropathology and histopathological findings
Autopsy findings from NiV-infected humans and animals demonstrate widespread vasculitis, necrotising lesions, syncytia in endothelial and neuronal tissues, micro-infarction, and evidence of viral antigen in brainstem medullary regions (Ng et al., 2004). Key features include:
5.4. Comparison with Hendra virus
While both NiV and HeV share similar mechanisms (Ephrin-B2/B3 receptor usage, endothelial/neuronal tropism), NiV tends to cause more frequent human-to-human transmission and has a broader geographic distribution. HeV outbreaks are primarily equine-to-human in Australia, and human-to-human spread is not documented. The relative severity of NiV may be linked to greater viral shedding in human respiratory secretions and wider host adaptability (Middleton et al., 2014).
In conclusion, the pathophysiology of NiV involves viral entry via ephrin receptors, endothelial injury, neuroinvasion, immunopathology and multisystem collapse, thereby explaining the clinical severity and rapid progression seen in many patients.
6. Clinical Manifestations
6.1. Incubation period
Reported incubation periods for NiV infection typically range from 5 to 14 days, although longer periods (up to 45 days) have been documented in rare instances (Luby et al., 2009; Wikipedia summary).
6.2. Early symptoms
The initial clinical presentation often resembles an influenza-like illness, with fever, headache, myalgia, vomiting, and sore throat. Patients may also experience fatigue and dizziness (Thakur & Bailey, 2019).
6.3. Progressive neurological and respiratory manifestations
In many cases, the disease rapidly progresses to involvement of the central nervous system (CNS) and/or the respiratory system:
6.4. Severe cases and chronic/relapsing manifestations
Severe NiV infection may lead to coma, multi-organ failure, and death within days of neurological onset (Wong et al., 2002). Importantly, relapsing or late-onset encephalitis may occur months to years after initial recovery; such cases present with altered mental status, seizures and high mortality (Hossain et al., 2008). Survivors may also experience long-term neurological sequelae including personality changes, residual seizures, and cognitive impairment (Neurologic signs review … 2020).
6.5. Differential diagnosis
Clinical presentation of NiV overlaps with other causes of viral encephalitis (e.g., Japanese encephalitis, Rabies, herpes encephalitis) and severe respiratory viral infections. Key differential features include epidemiological risk (bat exposure, date-palm sap, pig contact), rapid onset of encephalitis plus respiratory involvement, and known outbreak setting. Early recognition of NiV is critical to infection control and management. In summary, NiV infection presents a clinical spectrum from mild febrile illness to fulminant encephalitis and respiratory failure, with potential long-term consequences in survivors, underscoring the need for timely diagnosis and care.
7. Diagnosis
Fig no.2: Diagnostic / Laboratory assay image
7.1. Sample types and biosafety
Diagnosis of NiV infection requires collection of appropriate specimens under strict biosafety conditions (BSL-4 laboratories for virus isolation). Sample types include blood (serum/plasma), cerebrospinal fluid (CSF), throat/nasal swabs, urine and sometimes post-mortem tissue (CDC NiV factsheet). Proper transport, handling and PPE use (gloves, face shield, N95/FFP2, gown) are mandatory to avoid laboratory-acquired infections.
7.2. Diagnostic tests
7.3. Interpretation and timing
RT-PCR is most sensitive during early viraemic or respiratory shedding periods (first few days of symptoms). Serology becomes positive later in the course or during convalescence. Negative results do not exclude NiV if samples are collected late or inadequately. Close liaison with reference laboratories is essential for interpretation and confirmation.
7.4. Biosafety and reporting
Given the high-consequence nature of NiV, laboratories must adhere to WHO and CDC biosafety guidelines, including use of BSL-4 containment for virus culture and proper reporting of confirmed cases to public-health authorities (WHO, 2023). In summary, diagnosis of NiV infection relies on molecular and serologic assays conducted under rigorous biosafety frameworks, with emerging advances in rapid/field diagnostics offering promise for faster outbreak response.
8. Management and Therapeutic Strategies
8.1 Supportive and Symptomatic Treatment
Currently, there is no licensed specific antiviral therapy or vaccine for NiV. Clinical management revolves around intensive supportive care:
8.2 Experimental Antivirals
A number of candidate therapeutics have been evaluated pre-clinically or in few human cases:
Key pharmacological agents under investigation
Table 2. Key pharmacological agents for NiV under study.
|
Agent |
Mechanism of Action |
Status |
|
Ribavirin |
Broad-spectrum antiviral nucleotide analog |
Retrospective human use; mixed outcomes |
|
Favipiravir (T-705) |
Viral RNA polymerase inhibitor |
Animal model efficacy demonstrated; human trials pending |
|
Remdesivir |
Viral RNA polymerase inhibitor |
Preclinical data; no confirmed human trial yet |
|
m102.4 Monoclonal antibody |
Neutralises NiV-G glycoprotein |
Phase I/compassionate human use |
|
Fusion inhibitors / siRNA |
Inhibit viral membrane fusion or replication |
Early-stage research |
8.3 Vaccine Development
Efforts to develop NiV vaccines are progressing though no licenced product yet exists. Major platforms include:
In summary, while therapeutic and vaccine development for NiV is active and promising, current management remains supportive and highlights the urgency of further clinical research and global investment.
9. Future Directions and Challenges
While considerable progress has been made in understanding NiV, significant gaps and challenges remain, pointing to key directions for future research and policy.
9.1. Gaps in transmission ecology and host adaptation
Understanding the precise mechanisms of spill-over (bat to human, bat to livestock to human) remains incomplete. The influence of climate change, bat migration, deforestation and land-use change on NiV emergence warrants further investigation (Field et al., 2015). The potential for viral adaptation leading to sustained human-to-human transmission remains a major concern.
9.2. Limitations of diagnostics and treatment
Rapid point-of-care diagnostics for NiV are still not widely validated; improved field-deployable assays are needed, e

Kavita Kumbhar*, Sana Mulla, Dhanraj Jadge, The Nipah Virus: A Review of Epidemiology, Pathophysiology, and Management, Int. J. Med. Pharm. Sci., 2025, 1 (11), 216-227. https://doi.org/10.5281/zenodo.17757326
 10.5281/zenodo.17757326
10.5281/zenodo.17757326